|
|
|
А.Н. Стацкевич
Прежде чем начать рассказ о ТМОПВ, давайте окинем взглядом те гипотезы об упорядочивании воды вокруг белковых и других макромолекул, которые появились раньше.
История вопроса
Бертил Якобсон кратко в 1953, и более развернуто в 1955 году, предложил свою «гипотезу структуры воды», пытаясь объяснить диэлектрическое поведение водных растворов макромолекул.
Он писал: «Если на поверхности макромолекулы имеется много атомов кислорода и азота, причем их расположение идеально соответствует пространственной решетке воды, это производит сильный эффект упорядочивания и создает почти идеальную структуру, где каждая молекула воды соединена с четырьмя другими».
Образующиеся при этом гидратные оболочки Якобсон назвал «льдоподобными», чтобы акцентировать внимание на особенностях этой структуры, в отличие от структуры обычной воды, в которой имеются отдельные ассоциаты, каждая молекула в которых также может быть связана водородными связями с четырьмя другими, но доля таких ассоциатов в общем объеме невелика, а сами ассоциаты непрочны.
Двумя годами позже лауреат Нобелевской премии Альберт Сент-Дьёрдьи развил идею воды, подобной льду. Согласно «теории айсберга» Сент-Дьёрдьи, молекулы белков в клетках окружены кристаллами льда. Кроме того, идею льдоподобной воды вокруг белков также поддерживал Клотц.
Однако против всех теорий айсберга есть серьезное возражение.
Чамберс и Хейл, изучавшие замораживание переохлажденных мышц лягушки, обнаружили (а Миллер и Линг подтвердили), что в точках контакта кристаллов обычного льда с цитоплазмой переохлажденного мышечного волокна лягушки мгновенно образуются кристаллы льда. От них к концам мышечного волокна быстро вырастают длинные ледяные неветвящиеся лучи, прямые или искривленные в зависимости от того, вытянуто волокно или искривлено.
Этот пример показывает, что если бы в живой клетке постоянно существовали кристаллы льда, как утверждает теория айсберга, то ее переохлаждение сопровождалось бы спонтанным образованием льда вокруг этих кристаллов, как центров кристаллизации, и быстрым оледенением всей клетки, чего в реальности не наблюдается.
Тем не менее, и Якобсону, и Альберту Сент-Дьёрдьи — пусть их предположения и не оправдались — надо отдать должное за прекрасную аргументацию и интересные эксперименты, которые вновь привлекли внимание к столь важной проблеме, почти уже канувшей в лету после опытов Хилла с мочевиной.
ТМОПВ в клетке и ее мировое признание
ТМОПВ в клетке, неотъемлемая часть теории ассоциации-индукции (ТАИ), была опубликована спустя три года после выхода в свет собственно ТАИ. Однако лучше будет сначала рассказать о ТМОПВ, а потом уже — о собственно ТАИ, так как ТМОПВ, как и ТФЗЛ, касается статических свойств живых клеток. Динамические же, или индуктивные их свойства, которые всегда являются производными свойств статических, будут представлены позже при рассмотрении ТАИ.
Согласно ТМОПВ, вся или почти вся вода покоящейся клетки имеет отличную от обычной воды структуру динамичного характера, возникающую главным образом благодаря взаимодействию воды с сетью «полноразвернутых» белковых цепей, присущей всем клеткам.
Под полноразвернутым состоянием молекулы белка следует понимать такое ее состояние, при котором ни одна NH- или СО-группа полипептидной цепи не участвует в образовании иных водородных связей, кроме как с водой. Не следует путать полноразвернутую конформацию с «развернутой», как иногда называют в-складчатую конформацию белка. При некоторых допущениях полноразвернутую конформацию можно отождествить с так называемым «случайным клубком» и с тем, что Бунгенберг-де-Йонг называл линейной конформацией. Однако, по ТМОПВ, полноразвернутые белки и вода, которую они вокруг себя организуют, распределены в клетке отнюдь не беспорядочно.
Пептидный остов (цепь пептидных связей) белковых молекул представляет собой не что иное, как геометрически правильное чередование электрических диполей: отрицательно заряженных СО-групп (обозначаемых как N-центры, «отрицательные центры») и положительно заряженных NH-групп (обозначаемых как Р-центры, «положительные центры»), каждая из которых доступна клеточной воде, если белковая цепь полностью развернута, то есть не содержит элементов вторичной структуры. Одномерную (линейную) систему центров обозначим NP-системой. Двухмерную систему (поверхность) N- и Р-центров, расположенных в шахматном порядке, обозначим как NP-NP-систему.
Трехмерную сеть (или матрикс) подобных полноразвернутых белковых цепей можно назвать системой NP-NP-NP-типа, или NP-NP-NP-системой. Системой NO-NO-NO или РО-РО-РО-системой назовем трехмерную систему отрицательных (N) или положительных (Р) зарядов, правильно чередующихся с незаряженными (нейтральными) функциональными группами — О.
Наличие нейтральной группы между двумя ближайшими N- или Р-центрами оказывает сильное влияние на свойства матрикса потому, что от этих «вставок» зависит расстояние между соседними N- или Р-центрами.
Структура молекулы воды такова, что ее атомы располагаются в вершинах треугольника с атомом кислорода в вершине тупого угла. Каждая из связей кислорода с водородом представляет собой диполь с положительным (водород) и отрицательным (кислород) полюсами. Поскольку положительно заряженные полюса находятся относительно атома кислорода асимметрично, молекула воды имеет дипольный момент, равный в вакууме 1,85 дебая. Однако важно помнить, что эта величина не остается постоянной, она изменяется в зависимости от природы межмолекулярных контактов. Так, дипольный момент воды в жидком состоянии выше (2,9 дебая), чем в газообразном потому, что в присутствии других диполей или зарядов степень поляризации данного диполя обычно возрастает, он поляризуется.
Все N- и Р-центры полипептидного остова, расположенные вдоль него в геометрически правильном порядке, доступны воде, если белковая молекула полностью развернута. Взаимодействие этих центров с водой поляризует ее, делая водородные связи с другими молекулами более прочными, а NP-матрица определяет структуру первого слоя связанной воды таким образом, что дипольные моменты соседних молекул направлены в противоположные стороны (см. рис.). Этот первый слой, представляющий в свою очередь упорядоченную матрицу из полюсов диполей, адсорбирует следующий слой воды, дипольный момент которых при этом также возрастает. Формирование многослойной структуры будет продолжаться до образования мощной динамичной структуры поляризованных и ориентированных в пространстве молекул воды, «скрепленных друг с другом» более прочными водородными связями, чем в объемной воде. Структура, показанная на рис., — лишь стоп-кадр непрерывных изменений, способный создать ложное впечатление, что слои поляризованной воды предельно упорядочены и устойчивы.
|
|
Возникновение многослойной структуры поляризованной воды (кружки) под влиянием линейного матрикса из цепей полимера с чередующимися положительно и отрицательно заряженными фиксированными центрами (NP-система). Водные структуры, образовавшиеся на параллельно расположенных поверхностях (NP-NP-поверхность, не показана; ее плоскость перпендикулярна плоскости страницы) с чередующимися в шахматном порядке положительно и отрицательно заряженными фиксированными центрами (панель А) могут объединяться с образованием более толстых слоев поляризованной воды. На панели В показано, что многослойные структуры из поляризованной воды могут образовываться и на линейном матриксе, имеющем пробелы между заряженными группами (NO-система). В данном случае, на месте, где должен находиться положительный заряд, нет значимой для рассматриваемого процесса группы (NO-NO-поверхность не показана; ее плоскость перпендикулярна плоскости страницы). Совокупность плоских матриц образует трехмерную матрицу, заполненную ориентированными молекулами воды, организованными в многослойные структуры. Начало формированию такой структуры кладут взаимодействия фиксированных зарядов/диполей белка или другой макромолекулы с диполями воды, а сама водная сеть стабилизируется латеральными (внутрислойными) и радиальными (межслойными) водородными связями между самими молекулами воды. Первый слой адсорбированной воды, является матрицей для образования второго слоя и т. д. Поляризация молекул воды белком, передающаяся от слоя к слою, приводит к упрочению водородных связей и, соответственно, к увеличению устойчивости многослойной структуры связанной воды в целом (электростатические взаимодействия показаны стрелками). По этому принципу выстраивается кооперативная структура связанной воды (по Лингу). |
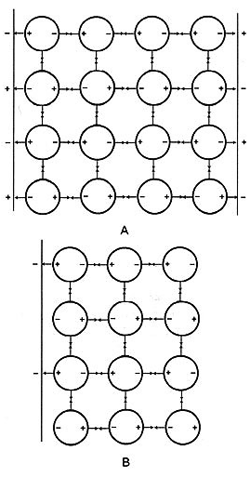
Необходимо лишний раз обратить внимание на понятие «поляризованная вода». Что оно означает в данном контексте? Поскольку молекула воды обладает дипольным моментом, то она уже поляризована. Однако принципиальным положением ТМОПВ является следующее: при взаимодействии с другими диполями или зарядами асимметрия зарядов в молекуле воды усиливается, ее дипольный момент возрастает. Именно эта дополнительная поляризация объясняет все свойства связанной воды в клетке и в модельных системах, отличающие ее от свойств объемной воды. Главным фактором, усиливающим дипольный момент воды, является полипептидный остов белков, и чем большая его часть доступна воде, тем больше воды будет модифицировано, будь то в клетке или в модельной системе. В полной мере это относится к любым другим макромолекулам, способным усиливать дипольный момент молекул воды и служить, как белки, матрицей для упорядоченной их адсорбции. Для простоты словоупотребления под «поляризованной водой» мы будем понимать воду с увеличенным дипольным моментом (наведенная поляризация), несколько превышающим дипольный момент молекул объемной воды.
В клетке (например, в мышечном волокне лягушки) толщина слоя поляризованной воды между соседними полноразвернутыми цепями белков составляет в среднем шесть молекул. Казалось бы, немного, однако на самом деле этого хватает, чтобы связать всю воду внутри мышечного волокна, превратив ее в многослойную динамичную структуру. А что значит «динамичная структура», спросите вы?
Тадж Махал не может сдвинуться со своего места — это пример статической структуры. Журавлиный клин — совсем другая структура: каждый ее элемент беспрестанно перемещается относительно некой средней позиции, но вместе они образуют всегда узнаваемую динамичную структуру. Точно так же, согласно ТМОПВ, ведет себя и вода в клетке.
Испытаем теперь эту теорию в эксперименте.
В такой сложной науке, как физиология клетки, создание и проверка новых теорий значительно упрощается, если уже на ранних этапах исследований имеется хорошая искусственная модель, демонстрирующая явления, предсказываемые теорией. Само существование убедительной искусственной модели, отвечающей требованиям теории, которая при этом, несмотря на свою простоту в сравнении с исследуемым живым прототипом, недвусмысленно воспроизводит то или иное его свойство — свидетельство истинности теории. И наоборот, если упорные поиски искусственной модели не увенчались успехом, это должно наводить на мысль об ошибочности теории — что, собственно, и имеет место, например, в случае с теорией мембранных потенциалов, о чем речь пойдет позже. История ТМОПВ в этом смысле была совершенно иной. Уже с первых шагов был найден целый ряд интересных модельных систем в ее поддержку.
По причинам, которые будут изложены позже, желатин, другие белки, денатурированные мочевиной и NaOH, а также линейные полимеры, несущие распределенные закономерным образом атомы кислорода или азота со свободной парой электронов — такие, как полиэтиленоксид (ПЭО), поливинил-пирролидон (ПВП), поливинилметиловый эфир (ПВМЭ) и полиэтиленимин (ПЭИ), — все являются полноразвернутыми соединениями. Благодаря этому, любой участок их молекулы доступен воде и легко с ней взаимодействует. Такие полностью открытые для взаимодействия с водой макромолекулы будем называть экстравертными моделями.
Молекула полиэтиленоксида (ПЭО) (—СН2СН2O—)n служит наглядным примером важности постулата ТМОПВ о значительном влиянии, которое оказывает расстояние между заряженными центрами полимера (атомами кислорода в данном случае) на характер его взаимодействия с водой. ПЭО, в молекуле которого атомы кислорода разделены двумя метиленовыми группами, хорошо растворим в воде и способен связывать и поляризовать ее на своей поверхности. Однако удаление одной метиленовой группы из повторяющегося звена полимера, либо добавление новых делает полимер нерастворимым в воде. Это свидетельствует об особой комплементарности ПЭО и воды, о том, что геометрия расположения атомов кислорода в цепи ПЭО играет важную роль в образовании водородных связей с молекулами воды, а между структурой связанной воды и положением атомов кислорода в полимере имеется стерическое соответствие.
Большинство же нативных глобулярных белков, а также белки, денатурированные SDS и n-пропанолом, относятся кинтровертным моделям, так как NH- и СО-группы их полипептидной цепи включены в б-спиральные или в-складчатые конформации и становятся недоступными воде.
По всему миру проводились исследования замечательных физико-химических свойств воды внутри клеток и клеточных моделей. Немало таких работ выполнено и в лаборатории Линга, но, в отличие от остальных исследователей, изучавших либо клетки, либо модели, группу Линга интересовала связь между ними.
У нас есть возможность провести сравнение результатов исследования клеток и модельных систем, полученных разными исследователями, по следующим направлениям (этот список мог бы быть значительно шире, если бы важность этой проблемы была очевидной для большего числа ученых):
- осмотическая активность
- набухание и сжатие
- понижение температуры замерзания воды
- поглощение пара при его парциальном давлении, близком к насыщению
- время вращательной корреляции ЯМР (τr)
- время диэлектрической релаксации Дебая (τD)
- коэффициент вращательной диффузии по данным квазиупругого рассеяния нейтронов
- растворимость веществ.
После сравнения результатов этих исследований стало очевидно, что вода, связанная экстравертными моделями, обладает точно теми же свойствами, что и вода в покоящейся клетке, тогда как у воды в интровертных моделях эти свойства отсутствуют или очень слабы — как это и следует из ТМОПВ. По понятной причине детальное обсуждение каждой из перечисленных характеристик не представляется возможным в рамках этой статьи. Однако мы постараемся ответить на два очень важных вопроса: сколько свободной воды имеется в типичной клетке? — к примеру, в мышечном волокне лягушки, — и какая доля воды в мышечном волокне поляризована и структурирована?
Отвечая на эти вопросы, мы фактически подвергнем строгой проверке саму ТМОПВ, согласно которой вся или почти вся вода внутри клеток, находящихся в состоянии покоя, включена в упорядоченные слои, а свободная вода практически отсутствует.
Ответ на вопрос о количестве свободной воды напрямую вытекает из сопоставления результатов двух простых экспериментов. Линг и Уолтон разработали метод удаления межклеточной жидкости центрифугированием: 4 минуты вращения в герметичном контейнере с ускорением 1000g приводит к удалению всей свободной воды из межклеточного пространства портняжной мышцы лягушки, не затрагивая воду внутри мышечных волокон. С помощью этого метода они попытались выжать всю воду из портняжной мышцы, разрезанной на фрагменты длиной 2 и 4 мм. Оказалось, что количество выделенной таким образом воды равно количеству воды во внеклеточном пространстве. Поскольку сверх этого количества из нарезанной мышцы выжать ничего не удалось (то есть внутриклеточная вода из разрезов не выходила), значит, свободной воды внутри самих мышечных волокон нет. Возможное возражение, что на поверхности разрезов быстро регенерировала мембрана, которая и заперла свободную воду во внутриклеточном пространстве, опровергается другим простым экспериментом, с очевидностью показавшим, что мышечные волокна лягушки не способны регенерировать плазматическую мембрану на усеченном конце. Итак, в мышце была обнаружена только одна фракция свободной воды — вода межклеточного пространства, свободное состояние которой ни у кого не вызывает сомнений.
Прежде же чем ответить на вопрос: какая часть клеточной воды организована в упорядоченные слои? - коротко остановимся на истории изучения сорбции газов.
Зависимость количества газа, сорбируемого твердым телом, от парциального давления данного газа часто представляет собой S-образною кривую. Вначале она идет круто вверх, затем следует плато, переходящее во второй крутой подъем. Прежде этот второй подъем поглощения в области предельных значений парциального давления объясняли капиллярной конденсацией. Но в 1934 году де Бур и Цвиккер предложили новое объяснение, основанное на их теории поляризации. Эта теория предполагает, что первый слой молекул газа адсорбируется на заряженные центры, расположенные в шахматном порядке на поверхности сорбента. При этом адсорбированные молекулы газа становятся электрическими диполями и, в свою очередь, поляризуют следующий слой молекул. Так адсорбируются все новые и новые слои молекул газа, образуя, в конце концов, многослойную структуру на поверхности твердого тела.
В 1952 году Брунауэр, Эмметт и Теллер подвергли теорию де Бура и Цвиккера резкой критике, указывая на то, что в результате электростатической поляризации может быть адсорбирован только один слой молекул, но не более того. Они предложили собственную теорию, известную по первым буквам их фамилий как теория БЭТ. Согласно теории БЭТ, первый слой молекул газа адсорбируется в соответствии с изотермой Ленгмюра. Остальные молекулы наслаиваются на первый вследствие обычной конденсации, поэтому их состояние напоминает состояние жидкого газа.
Однако Брунауэр и его коллеги при этом ясно дали понять, что их критика теории де Бура и Цвиккера касается лишь адсорбции газов, близких по свойствам к благородным, — то есть не обладающих собственным дипольным моментом. А механизмы адсорбции газов, чьи молекулы обладают значительным собственным дипольным моментом (именно их исследовал Бредли), под их критику не подпадают (напомним, что молекулы воды тоже обладают собственным дипольным моментом).
В 1965 году, представляя ТМОПВ применительно к клетке, Линг отметил, что теория БЭТ способна удовлетворительно объяснить лишь ограниченный круг сорбционных явлений, таких как поглощение воды овечьей шерстью и коллагеном при очень низком парциальном давлении водяного пара. Данные же о поглощении воды различными белками и полипептидами в широком диапазоне парциального давления водяного пара, полученные Меллоном, Корном и Гувером, соответствуют изотерме Бредли. Хотя Брунауэр и его коллеги не нашли аргументов против взглядов Бредли на сорбцию молекул газов с собственным дипольным моментом, они не стали углубляться в эту проблему. Возможность многослойной адсорбции молекул воды, обладающих значительным собственным дипольным моментом, определяется сочетанием двух важных факторов — поляризацией и ориентацией ее молекул матрицей N- и Р-центров. Именно благодаря этой поляризации-ориентации исходный дипольный момент воды может несколько увеличиться. В результате адсорбированные молекулы воды ориентируются таким образом, что способны в свою очередь поляризовать и ориентировать в пространстве следующий слой молекул воды, а за ним еще и еще. В идеальных условиях — при достаточной плотности заряда на N- и Р-центрах, и при температуре абсолютного нуля — энергия взаимодействия молекул воды в поляризованных и структурированных слоях будет оставаться неизменной на любом удалении от поляризующей поверхности. Хотя в реальности эти условия недостижимы, распространение эффекта поляризации и ориентации на сверхдлинные расстояния теоретически возможно, о чем уже есть сообщения (правда, ТМОПВ нет необходимости выдвигать столь суровые температурные и другие требования для того, чтобы обеспечить свою работоспособность).
Как уже говорилось, изотерма адсорбции Бредли имеет дело с многослойной адсорбцией, причем каждый слой представляет собой мозаику из «кусочков» правильной формы — молекул, сориентированных в определенном направлении в результате взаимодействий как с соседями по своему слою, так и с соседями из других слоев; такая модель работает, если адсорбированные молекулы имеют собственный дипольный момент, а на сорбционной поверхности имеются фиксированные заряды/диполи, расположенные на ней в правильном порядке (например, как клеточки шахматной доски). Согласно ТМОПВ, в нормальных клетках, находящихся в состоянии покоя, большая часть воды, если не вся, адсорбирована по Бредли (как и вода в экстравертных моделях на основе желатина или других сходных по свойствам макромолекул).
Для проверки этого предположения Линг и Негенданк исследовали в стерильных условиях при 25°С равновесную сорбцию воды тонкими пучками мышечных волокон, выделенных из изолированных портняжных мышц лягушки. Время установления диффузионного равновесия оказалось равным 7-8 суткам. Давление пара варьировали почти от нуля (0,043) до почти полного насыщения (0,996) с помощью растворов NaCl и серной кислоты разных концентраций. Полученные данные доказывают, что состояние 95% воды в мышечных волокнах соответствует, как и предсказывала ТМОПВ, сорбционной модели Бредли. Остальные 5% также связаны, но свойства этой фракции отвечают требованиям однослойной адсорбции Ленгмюра; эта вода характеризуется прочной связью с адсорбентом — очевидно, полярными боковыми цепями клеточных белков.
Данные Юлиуса Катца о сорбции воды желатином, опубликованные в 1919 году, также охватывают весь диапазон влажности. Сделанный Лингом анализ этих данных показал, что и они подчиняются изотерме Бредли.
В предыдущей статье мы рассказывали об исследованиях Енё Эрнста, Иозефа Тидьи с сотрудниками равновесной сорбции воды портняжными мышцами лягушки. Большая часть их экспериментальных данных была получена на мышцах с содержанием воды не более 10% от нормы и характеризует поэтому только воду, прочно связанную белками.
Бегло рассмотрев основные экспериментальные данные в пользу ТМОПВ, перейдем теперь к теоретическим и практическим следствиям, вытекающим из ее принципиальных положений.
Следствия ТМОПВ для теории и практики и их подтверждение
Изобретение магнитно-резонансной томографии (МРТ)
Молекулы поляризованной воды, образующие многослойную динамичную структуру, постоянно перемещаются, меняются друг с другом местами и т.д. Но если бы нам удалось сделать стоп-кадр, то оказалось бы, что взаимодействие большинства молекул с соседями (а также, опосредованно, с группами пептидной связи — СО и NH, — организующими и поддерживающими всю эту структуру) сильнее и устойчивее, чем в обычной воде именно благодаря поляризации — увеличению дипольного момента молекул воды. Существование такой сети межмолекулярных связей ограничивает и замедляет как поступательное, так и вращательное движение молекул воды. Ограничение свободы перемещения молекул воды и входящих в их состав атомов водорода (или протонов) в многослойных структурах — одно из важнейших следствий ТМОПВ.
Фриман Коуп, получивший физическое образование в Гарварде и медицинское в Медицинской школе Джонса Хопкинса, одним из первых осознал, что, если утверждение ТМОПВ насчет ограниченной подвижности молекул воды в клетках истинно, то это можно обнаружить при помощи такого физического прибора как спектрометр ядерного магнитного резонанса (ЯМР).
Этот прибор позволяет измерить время вращательной корреляции (фr) атомов или протонов водорода в молекулах воды, или время их релаксации Т1 и Т2. Не вдаваясь в детали, скажем, что фиксация молекул воды (а, следовательно, и ядер атомов водорода) и ориентация их в пространстве облегчает размагничивание протонов, известное как релаксация. Согласно ТМОПВ, протоны клеточной воды менее мобильны, чем протоны обычной воды, поэтому время T1 и Т2 для них должно быть короче. Вскоре Коуп подтвердил эту догадку. Впрочем, не он один.
Одновременно с ним другой молодой физиолог Карлтон Хейзелвуд, также из Медицинской школы Джонса Хопкинса, сделал со своими коллегами это же открытие. Затем на сцену вышел третий молодой ученый по имени Реймонд Дамадьян. Взяв на время нужный ему прибор и выпросив у Института Слоуна и Кеттеринга несколько крыс, больных раком, он сделал выдающееся открытие.
Дамадьян показал, что вода в разных тканях отличается по временным параметрам Т1 и Т2. А в злокачественных опухолях Т1 и Т2 протонов воды значительно больше, чем в клетках тканей, из которой эти опухоли произошли. Так, протоны воды в опухоли печени — гепатоме — имеют гораздо большие T1 и Т2, чем протоны в нормальной печеночной ткани.
Дамадьян сразу понял, что это различие в Т1 и Т2 может лечь в основу нового медицинского прибора, которого еще не существовало, и который позволит обнаруживать злокачественные опухоли, не нанося никакого вреда организму больного.
Таким образом, сбылись пророческие слова Альберта Сент-Дьёрдьи о том, что раковые клетки имеют «меньше водных структур», сказанные им в 1957 году в примечании к одной из его работ. Его мысль о меньшем количестве структурированной воды в опухолевых клетках вполне согласуется с ТМОПВ. Однако дальнейшие исследования показали, что увеличение доли свободной воды — лишь одна из причин увеличения Т1 и Т2 в опухолевых тканях.
Еще не успели стихнуть насмешки критиков, когда Дамадьян с двумя аспирантами — Ларри Минкоффом и Майклом Голдсмитом — запустили девятый по величине сверхпроводящий электромагнит в мире и в мгновение ока соорудили на его основе первый аппарат ЯМР, окрещенный «Непобедимым» и ныне стоящий в одном ряду с другими историческими изобретениями человечества.
А 15 июля 1988 года Дамадьян вместе с Полом Лотербуром, предложившим более совершенную технологию получения ЯМР-изображения, удостоились от президента Рональда Рейгана Национальной технологической премии за изобретение нового метода исследования, ныне известного как магнитно-резонансная томография, или МРТ. Дамадьяну, кроме того, 12 февраля 1989 года была посвящена церемония посвящения его в изобретатели Национального зала славы изобретателей США.
Но лучше всего мечтательную и благородную натуру этого человека характеризует следующее письмо, которое он написал Гильберту Лингу 9 ноября 1977 года:
«Утром 3 июля, в 4:45 утра... мы в великом ликовании получили первое в мире МРТ-изображение тела живого человека. Это достижение стало возможным благодаря современным концепциям биофизики водных растворов, великим пионером в которой стали Вы с Вашим классическим трудом — теорией ассоциации-индукции».
Немногие написали бы такое письмо другу (а тем более — неизбежному сопернику в науке). Но это благородство претворилось в еще нечто более важное — в поступок, когда он и созданная им компания Fonar, производящая аппараты МРТ, предложили поддержку и пристанище Лингу и его коллегам Маргарет Оксенфельд и Чжэнь-дон Чэню после того, как лаборатория Линга была закрыта на пике своей продуктивности по инициативе его научных оппонентов и примкнувшего к ним Национального института здоровья США. В 1988 году Лингу с сотрудниками пришлось распрощаться с Пенсильванским госпиталем в Филадельфии, первой больницей в истории США и пристанищем для группы Линга и их исследований на протяжении 27 лет.
Вскоре молодой компании Fonar пришлось изо всех сил биться за выживание с такими гигантами мировой индустрии, как General Electric, Siemens, Phillips, Hitachi, Toshiba, не говоря уже о множестве других компаний поменьше. Однако в США сложилась самая совершенная на сегодня патентная система в мире, которая не только защищает и вознаграждает как мелких, так и крупных изобретателей, но и, что не менее важно, обеспечивает, благодаря своей открытости, широкий доступ к своим патентам по всему миру, что служит интересам научно-технического прогресса. В 1997 году Верховный суд США, утвердив решения суда низшей инстанции и Апелляционного суда, предписал компании General Electric, гиганту из гигантов индустрии, выплатить компании Fonar 128,7 миллионов долларов за нарушение ее патента на новаторский метод диагностики рака, а также еще одного патента. Правда, немалая часть этих денег ушла адвокатской конторе «Robins, Kaplan, Miller and Ciresi», отстаивавшей иск Fonar.
Все то, что ныне нам известно о клеточной воде, стоило больших жертв и другим ученым. Один из первых исследователей клеточной воды методом ЯМР, Карлтон Хейзелвуд пережил гонения и суровые лишения за непоколебимость своих научных взглядов. Джим Клег, среди прочих своих научных достижений, подтвердил ограниченную подвижность клеточной воды, исследуя ее диэлектрические свойства в ультравысокочастотном электрическом поле. Бад Роршах стал инициатором исследований коэффициентов вращательной диффузии методом квазиупругого рассеяния нейтронов. Трагедией стало самоубийство Фримана Коупа в 1982 году, после того, как была прекращена финансовая поддержка его научной работы.
Подготовлена с использованием материалов книги Гильберта Линга "Физическая теория живой клетки"
А.Н. Стацкевич, Осмысление пройденного 4. Теория многослойной организации поляризованной воды в клетке (ТМОПВ) // «Академия Тринитаризма», М.,
|
|