|
|
|
А.Н. Стацкевич
(По материалам Барбараш А.Н. Основы стереогенетики)
Важным доводом в пользу волнового управления генами эукариот является строгая пространственная организация эукариотических геномов.
У безъядерных клеток (прокариот) замкнутая в кольцо нежесткая спираль ДНК свободно плавает в протоплазме и либо вовсе ни к чему не прикреплена, либо прикреплена к оболочке клетки в небольшом числе точек (от 1 до 100). Говорить о стабильности или закономерности пространственного расположения генов здесь не приходится. При ядерной же организации генома основная часть ДНК содержится в гетерохроматине, который имеет вид жестко фиксированной пространственной структуры, во множестве точек прикрепленной к ядерной оболочке и к скелетной конструкции, называемой ядерным матриксом.
Ядерный матрикс состоит из двух компонентов:
- немембранной части ядерной оболочки (ламины), связанной с поровыми комплексами оболочки, и
- внутриядерной сетчатой структуры из фибрилл и гранул [Збарский, 1988].
Ядерный матрикс построен из кислых негистоновых белков, содержит несколько процентов РНК, ДНК и липидов. Небольшое количество ДНК, обнаруживаемое в выделенном ядерном матриксе, вероятно, представляет собой прочно прикрепившиеся участки удаленных хромосом.
Кроме компактного гетерохроматина в ядре существует разрыхленная, активная фракция, называемая эухроматином. В ней строгое пространственное положение занимают только гены, еще не подвергшиеся активированию. Разрыхление хроматина, переводящее ген в активное состояние, делает его пространственные координаты менее определенными, превращает этот короткий участок хромосомы в свободно плавающую петлю ДНК. После прекращения акустических колебаний, самосборка хроматина снова возвращает ген в заданную микроскопическую зону пространства ядра, где он будет ожидать нового активирования.
Пример прокариот показывает, что гены способны нормально функционировать и без закрепления в определенных точках пространства. В то же время, в ядрах всех эукариот (а это около двух миллионов видов) гены зафиксированы, из чего видно, что фиксация, жестко контролируемая естественным отбором, стала жизненно необходимым условием существования организмов. Благодаря фиксации реализуются определенные закономерности пространственного расположения генов внутри ядер — биологам известен ряд таких закономерностей.
Выяснено, что хромосомы располагаются в ядре в строго определенном порядке. Для этого Природой выработан остроумный механизм, основанный, в частности, на однозначном взаимном расположении хромосом, благодаря определенным длинам их плеч. Если отбросить экзотические варианты геномов, требующие отдельного рассмотрения (например, геномы с мини-хромосомами), то этот механизм, открытый новосибирским биологом А. И. Щаповой в 1971 году, выглядит следующим образом.
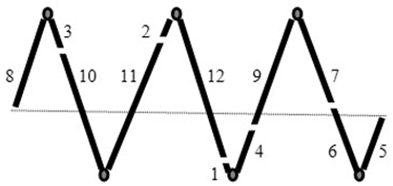
Рис. 1: Пример задания взаимного расположения хромосом в ядре через неповторяющиеся длины их плеч (кружками обозначены центромеры, цифрами — номера плеч хромосом в порядке увеличения их длин; плечи 5 и 8 соединяются между собой, подобно другим парам плеч).
При распределении плеч хромосом какого-либо генома (например, пшеницы или человека) по порядку возрастания их длин, этот ряд образует более или менее равномерную лесенку, где не встречается двух плеч одинаковой длины. В ходе клеточного деления, хромосомы дочерних клеток (в ранней стадии профазы) строго определенным образом соединяются концами (теломерами), образуя замкнутую цепочку. Поскольку в районе центромер хромосомы при этом изламываются под острым углом, цепочка приобретает вид замкнутой змейки, как это схематически изображено на рис. 1. Каждый гаплоидный набор хромосом образует самостоятельную замкнутую змейку. Гомологичные хромосомы двух гаплоидных геномов располагаются рядом, бок о бок.
Так как лесенка плеч хромосом более или менее равномерна, то при соответствующем соединении плеч между собой становится возможным построение такой змейки, у которой все расстояния между центромерами (вершинами змейки) одинаковы. Но при наличии в гаплоидном геноме каждой длины плеча в одном экземпляре, одинаковое расстояние между центромерами оказывается возможным лишь в единственном варианте взаимного расположения хромосом в змейке [Щапова, Баутина, 1975; Щапова, Потапова, 1984]. Позже было установлено, что теломеры хромосом содержат строго индивидуальные для каждой хромосомы тандемные повторы, которые, вероятно, и образуют так называемые «липкие концы», быстро и однозначно соединяющие нужные теломеры в процессе формирования змейки.
Объединение четных и нечетных вершин змейки, соответственно, у одного и у другого полюса формирующегося ядра позволяет расположить пары плеч вдоль меридианов, как показано на следующем рис. 2. Не исключено, что временному соединению центромер в два «узла» у полюсов ядра помогают индивидуальные особенности тандемных повторов центромерных участков хромосом. Весь этот механизм служит для того, чтобы отрезки змейки заняли четкое меридиональное положение перед последующими операциями разворачивания пространственной структуры генома.
Рис. 2: Схема расположения хромосом в ранней профазе митоза у пшеницы-однозернянки Triticum monococcum [Щапова, 1971]
На дальнейших стадиях формирования ядра (в поздней стадии профазы) происходит ряд перестроек, в том числе разъединяются концы хромосом, все центромеры направляются к одному полюсу ядра, а теломеры — к другому, каждая из хромосом начинает по-своему изгибаться, принимая определенную конфигурацию. Эта конфигурация и фиксируется, в конце концов, прикреплением ДНК к ядерному матриксу [Hancock, Hughes, 1982] и оболочке ядра.
Результатом временного соединения хромосом в замкнутую змейку оказывается одинаковый для всех клеток организма порядок (очередность) взаимного расположения хромосом (и их плеч) вдоль экватора ядра.
Кроме закономерного расположения хромосом в ядре, есть данные и о закономерном размещении генов на хромосомах. Лима-де-Фария ввел представление о центромеро-теломерном поле как о своеобразной шкале отсчета координат гена на хромосоме (аналогично «географической широте» по отношению к полюсам ядра). Исходя из таких представлений, он классифицировал различные гены как центроны, медоны и телоны, в зависимости от того, расположены ли они около центромеров, в медиальной (средней) части плеч или около теломеров. Например, положение генов 28S и 18S, кодирующих рибосомную РНК (область ядрышкового организатора), проверенное для 100 видов — от водорослей до человека, — соблюдалось настолько строго, что его можно было предсказывать исходя из простого уравнения. Аналогичные закономерности показаны для групп крупных хромомеров, для генов, чувствительных к холоду, и в других случаях [Лима-де-Фария, 1981].
Закономерное расположение генов на хромосомах и хромосом в ядре дополняется принятием хромосомами определенных конфигураций в пространстве ядра. В каждом типе дифференцированных клеток хромосомы изгибаются внутри ядра строго определенным образом. Эти три закономерности в итоге выливаются в общее закономерное пространственное расположение генов в интерфазных ядрах. Но из-за разных изгибов хромосом внутри ядер клеток разных типов, даже в одном организме, одинаковое расположение генов в ядре бывает только у однотипных клеток. Другими словами, при неизменном положении генов на хромосомах, их пространственное расположение в ядре закономерно изменяется в ходе дифференцировки клеток [Щапова, Баутина, 1975; Steffensen, 1984].
Яркий пример закономерной пространственной организации показывают три наиболее крупных гена рибосомальных РНК (рРНК) — гены 28S, 18S и 5,8S. Ничем не отличаясь у прокариот от других генов, эти гены, попав в ядро, заняли во всех клетках строго определенное пространственное положение в центральной зоне ядра, не допуская сюда никакие другие гены. Упомянутые гены рРНК не покидают центральной зоны ядра и при дифференцировке клеток, когда пространственные конфигурации хромосом кардинально перестраиваются.
Можно ли объяснить строгие закономерности пространственного расположения и фиксацию генов внутри ядер всех эукариот, в сочетании с прекрасным функционированием свободно плавающих нуклеоидов прокариот? Вспомним, что ТДС не дает такого объяснения. Что, опять пытаться объяснять ситуацию случайностью и просто удивительным совпадением фактов с выводами КСГ и требованиями оптической проекции?
Известны также закономерности, не относящиеся непосредственно к пространственному расположению генов в ядре, но, тем не менее, указывающие на небезразличное пространственное поведение хромосом, в отличие от ДНК прокариот. Например, у хромосом гибридов Hordeum*Secale на стадии метафазной пластинки замечено разделение по отношению к центру клетки; во всех исследованных случаях хромосомы Hordeum оказывались ближе к центру, чем хромосомы Secale [Беннет, 1986].
Более того, оказалось, что некие закономерности разделения хромосом в метафазной пластинке можно заметить не только у гибридных геномов, но и в диплоидных геномах гомозиготных линий, где гомологичные хромосомы практически одинаковы! В этом случае хромосомы разных гаплоидных наборов стремятся расположиться по разные стороны от воображаемой прямой линии, делящей метафазную пластинку надвое.
Еще одна закономерность заключается в том, что одинаковые хромосомы двух гаплоидных наборов расходятся дальше друг от друга, чем разные. Это противоречит предположению, что у гибридов гаплоидные наборы хромосом отдаляются друг от друга из-за своего несходства. Здесь проглядывает какая-то более сложная зависимость.
Принципиальная важность вопросов пространственной организации геномов эукариот видна и при изучении эволюции. При переходе от прокариот к эукариотам центр тяжести эволюционных изменений переместился с кодов белков на пространственное расположение генов в ядрах. Обнаружилось, что скорость эволюции крупных таксонов коррелирует со скоростью перестроек структуры ядра [Nei, 1975].
За последние десятилетия существенно изменились взгляды биологов на возможную скорость изменения геномов. Изменения, весомо меняющие свойства популяции, ранее воспринимались, в основном, как элементы эволюционного процесса, отмеряемого по шкале в тысячи или миллионы лет. Никто не видел в геноме одну из регуляторных систем, сравнительно быстро приспосабливающих популяцию к меняющимся условиям обитания. Тем более, никто не подозревал, что геномы могут с высокой скоростью изменять свойства популяции как в одном, так и в противоположном направлениях.
Но именно такие способности генетических систем обнаружились при исследованиях мобильных диспергированных генов — МДГ.
Строго говоря, первооткрывателям не следовало называть эти элементы генами, поскольку МДГ, по-видимому, не содержат кодов для синтеза каких-то молекул. Большинство исследований выявляет в них свойства групп коротких неинформативных повторов.
Основная масса исследований МДГ проводилась на дрозофилах, хотя использовались и другие организмы, например, дрожжи.
Выяснилось, что при навязывании популяции экстремальных условий существования, в ее геномах быстро перемещаются некоторые последовательности ДНК, и синхронно происходит общее повышение жизнеспособности популяции. При создании для дрозофил тяжелых условий существования, усиливших отбор на половую активность и жизнеспособность, в популяции изменилась локализация одного из семейств МДГ. Число сайтов их локализации увеличилось в разных экспериментах от 14–15 до 21–25. В некоторых экспериментах наблюдалось скачкообразное перемещение и увеличение числа сайтов локализации за одно поколение с одновременным скачком жизнеспособности и конкурентоспособности. Характерный для измененной популяции геном практически полностью стабилизировался за четыре поколения [Беляева и др., 1981].
«Исходные линии четко различаются по числу и локализации в хромосомах копий МДГ. Они сохраняют высокую стабильность на протяжении десятков и сотен поколений. Частоты перемещений МДГ-элементов близки по грубым оценкам к 10–4. Направленный посемейный отбор на повышение приспособительных свойств приводит к выделению отдельных редких семей, которые превосходят остальные семьи по селектируемым признакам … У них, как оказалось, происходит реконструкция генома, выражающаяся в изменении числа и мест локализации МДГ-элементов. Копии МДГ появляются в новых сайтах, их число, как правило, увеличивается. Копии из старых сайтов обычно исчезают … Процесс перестройки генома такого рода захватывает сразу множество генетических элементов. Копии МДГ в новых сайтах часто появляются кооперативно, целыми блоками».
Центральным моментом исследований стало обнаружение в геноме горячих точек-мишеней, куда с высокой вероятностью перемещаются копии МДГ при селекции на повышение приспособительных свойств… Приобретение… высокого уровня приспособленности сопровождается появлением 5–8 новых сайтов локализации копий… МДГ в горячих точках… Возвратная селекция в минус-направлении по адаптивно важным свойствам сопровождается исчезновением копий МДГ из горячих точек». [Кайданов, 1991]
Воспроизводимость результатов и совпадение перемещений МДГ в родственных экспериментах продемонстрировали их закономерность. С позиций ТДС, подобная связь перестроек генома с изменением свойств организмов совершенно непонятна.
Вероятно, в перемещениях МДГ виден не сам процесс ускоренной эволюции дрозофил (невозможно, чтобы эволюция могла протекать с такой высокой скоростью), а быстрое повторение ранее пройденного пути, не раз использованного для выживания популяции в тяжелых условиях обитания и сохраненного геномом в виде системы «горячих точек».
Как отмечалось ранее, перемещения МДГ, вероятно, открывают собой более широкий, пока еще мало изученный класс быстрых генетических регуляций, очень существенных для биологической теории и практики. Надо думать, его следовало бы обозначить специальным термином, например, «быстрая адаптация генома» — БАГ.
В исследованиях МДГ для нас важно то, что быстрые изменения свойств новых поколений вызываются пространственными перегруппировками генетического материала, а не появлением на хромосомах генов с новыми свойствами. Важен сам факт, что разному пространственному расположению МДГ соответствуют разные свойства организма. Феномен МДГ лишний раз показал, что при переходе от прокариот к клеткам с ядрами пространственная организация генома стала играть исключительно важную роль. Связь изменений на уровне организма с перестройками внутри ядра свидетельствует о существовании некоего соответствия между пространственной структурой генома и структурой организма в целом
Это говорит в пользу какого-то осовремененного варианта преформизма, в пользу существования в клетках «плана строения» организма, с которым связан организм в целом. С позиций КСГ, здесь мы видим проявление оптической проекционной связи организма с хроматином ядер.
А.Н. Стацкевич, Что и как управляет морфогенезом? Часть 9. О внутриядерной перестройке генома // «Академия Тринитаризма», М.,
|
|