|
|
|
А.Б. Савинов
Проблема «нового синтеза» или создания принципиально иной эволюционной парадигмы в противовес синтетической теории эволюции (СТЭ) поставлена давно, но до настоящего времени не решена. Современная эволюционная парадигма не должна формироваться путем синтеза положений, только альтернативных СТЭ. Новая парадигма эволюции может быть создана, во-первых, на основе диалектико-материалистического и системно-кибернетического подходов к биологическим объектам и явлениям, во-вторых, путем интеграции рациональных элементов СТЭ и альтернативных взглядов.
Problem of the new evolutionary paradigm (philosophical, system and general-biological aspects)
Savinov A.B.
The problem of ""» new synthesis ""» or creation essentially other evolutionary paradigm in a counterbalance of the synthetic theory of evolution (S
ТE) is put for a long time, but till now is not solved. The modern evolutionary paradigm should not be formed by synthesis of the positions only alternative SТE. The new paradigm of evolution can be created, first, on the basis of dialectics-materialistic and system-cybernetic approaches to biological objects and the phenomena, secondly, by integration of rational elements SТE and alternative sights.
Проблема «нового синтеза» или создания принципиально иной эволюционной парадигмы (Красилов, 1984, 1986; Мейен, 1984; Хохряков, 1984; Назаров, 1991) в противовес синтетической теории эволюции (СТЭ) поставлена давно, но до настоящего времени не решена (Гродницкий, 2002, Назаров, 2005). Это создает трудности методологического характера в эволюционно-биологических исследованиях и для процесса естественно-научного образования в школе и вузах (особенно на биологических факультетах университетов). Такую ситуацию противники материалистического мировоззрения (Струминский, 1995; Хлебосолов, 2004; Карпачевский и др., 2006) пытаются использовать для возрождения креационизма и теологизации науки.
Доминирующая в отечественной биологии СТЭ (Иорданский, 2001; Яблоков, Юсуфов, 2004; Северцов, 2005) подвергается критике, в ряде случаев справедливой, хотя взгляды ее противников не всегда последовательны. Так, обычно их главные возражения направлены против базовых концепций СТЭ о мутационном процессе и естественном отборе как ведущих эволюционных факторах. В противовес выдвигаются, во-первых, положения о ведущей роли в филогенезе факторов внешней среды (экосистемных, биосферных, космических); во-вторых, концепция эпигенетики как нового направления биологии (в том числе, эволюционной), но трактуемого неоднозначно (Гродницкий, 2002; Назаров, 2005; Корочкин, 2006; Чадов, 2006; Чураев, 2006; Шишкин, 2006) и «насыщаемого» иногда сложными или малопонятными абстракциями: «эпиген», «ячейка функциональной наследственной памяти» (Чураев, 2006); в-третьих, учение о симбиогенезе, противопоставляемое концепции борьбы за существование.
По-видимому, кризисное состояние эволюционного учения в наши дни в наибольшей степени обусловлено произвольным толкованием явлений, объединяемых неоднозначно трактуемым термином «эпигенетика» (с помощью которого в 40-х годах прошлого века К. Уоддингтон попытался всего лишь обособить исследования онтогенетических взаимодействий между генами и их продуктами, слагающими фенотип). Об этом свидетельствуют «эпигенетические» статьи и книги по теории эволюции последних лет. Так, например, Д.Л. Гродницкий (2002, с. 62, 63) полагает, что «эпигенетика — синоним биологии развития или эмбриологии на клеточном уровне», при этом «эпигенетическая система есть нечто, находящееся над фенотипом и генотипом и определяющее их изменение в ходе эволюции», а «эпигенетическая теория (эволюции — А.С.) рассматривает эволюцию как процесс преобразования онтогенеза под влиянием изменений, происходящих в окружающей среде» за счет генетической ассимиляции морфозов.
Иные, неоднозначные толкования эпигенетических явлений и их эволюционной роли обнаруживаются в работах ведущих отечественных генетиков. Своеобразным апогеем противоречивости во взглядах на эпигенетику явилась подборка статей в тематическом выпуске журнала «Генетика» (2006, № 9), целиком посвященном выяснению предмета, целей, задач эпигенетики и обсуждению результатов эпигенетических исследований, выполненных за последние годы мировым научным сообществом.
Журнал открывает статья Л.И. Корочкина (2006) «Что такое эпигенетика», которая, возможно, по замыслу редколлегии должна была стать «программной». В этой статье автор попытался ответить на вопрос, поставленный в названии статьи, объяснить принципиальное своеобразие, суть эпигенетических процессов, резко критикуя альтернативные взгляды на эпигенетику, в том числе отраженные в некоторых других статьях данного журнала. В частности, было отмечено, что в настоящее время «об эпигенетике много говорят и много пишут, часто не отдавая себе отчета, в чем, собственно смысл этого понятия», «в результате формируются крайне расплывчатые представления как о самой эпигенетике, так и о смежных с нею областях знания»; при этом нередко сама эпигенетика, этот термин «представляется как панацея от всех бед, как слово, разрешающее все противоречия и снимающее все проблемы в биологии и генетике развития, в физиологии и даже в эволюционной концепции», «требующее смены парадигмы чуть ли не всей биологии» (Корочкин, 2006, с. 1156). «Даже некоторые наши достаточно квалифицированные биологи путают запись наследственной информации и ее реализацию, утверждая, будто носителем наследственности является и ядерная мембрана, и клеточные белки, и прочие вещи, нужные для формирования признака», «нужные, но только для реализации того, что записано в ДНК как в материальной носительнице наследственности», — отмечает Л.И. Корочкин (2006, с. 1162) и сетует: «Неужели это так трудно понять! И не сеять смуту там, где нет для этого никаких оснований!». Он подтверждает, что «эпигенетический статус клетки передается дочерним производным при ее делении», но это не означает наследования приобретенных признаков, под которым «подразумевается ... передача из соматических клеток неких качеств, приобретенных путем, например, упражнений, в геном половых клеток и закрепление их там с последующей передачей потомству» (Корочкин, 2006, с. 1161).
Судя по всему, эти, весьма категоричные, критические замечания были высказаны в том числе и в адрес статей Б.Ф. Чадова (2006) «Новый этап в развитии генетики и термин «эпигенетика»» и Р.Н. Чураева (2006) «Эпигенетика: генные и эпигенные сети в онто- и филогенезе», материалы которых концептуально, идейно противоположны содержанию статьи Л.И. Корочкина (2006). Так, по мнению Б.Ф. Чадова (2006, с. 1262), на современном этапе развития генетики «в качестве генов будут фигурировать не ДНК и ее участки, а информационные продукты, образующиеся на ДНК», в связи с чем необходимо переименовать ДНК в прогены, а ее информационные продукты — в гены. Б.Ф. Чадов аргументирует свою позицию, во-первых, тем, что ДНК и ее участки представляют вещество наследственности, но их нельзя считать генами, поскольку последними традиционно называются дискретные материальные единицы, передающиеся с гаметами от родителей потомкам. Молекулы ДНК и хромосомы, хотя и передаются с гаметами, но материально едины, т.е. дискретными не являются. Во-вторых, указывает Б.Ф. Чадов, генетическая информация, заключенная в родительской ДНК, не реализуется и не вовлекается в «информационный поток» без специфических дискретных переносчиков — РНК, регуляторных белков и мобильной ДНК, ведущих себя независимо. Поэтому, полагает Б.Ф. Чадов, именно эти переносчики следует называть генами, а исходную ДНК и отдельные ее участки — прогенами, входящими в состав прогенома. В соответствии с таким подходом, «наследственность передается в виде прогенов, а реализуется с помощью генов», тогда «с точки зрения эпигенетики информационные продукты, названные ... генами, это — эпигены, а ДНК и ее районы — по-прежнему гены» (Чадов, 2006, с. 1263). Представляется, что предлагаемое переименование генов в прогены нецелесообразно, хотя бы в силу неизбежной терминологической путаницы.
Обратимся к классикам. По определению одного из родоначальников молекулярной биологии и первооткрывателей структуры ДНК Дж. Уотсона (1967, с. 227) «ген — это дискретная область хромосомы, ответственная за образование определенного клеточного продукта; он состоит из ряда линейно расположенных единиц, потенциально способных к изменению (мутирующих участков); каждый такой участок может существовать в нескольких альтернативных формах, и между разными участками может происходить кроссинговер». По мнению М.Е. Лобашева и соавторов (1979), ген — это функционально неделимая единица наследственности (наследственной информации), делимая при кроссинговере, но не являющаяся единицей мутирования, т.е. ген — это лишь задаток, дающий возможность развития отдельного признака (свойства), касается ли это структуры белковой молекулы или элементарной реакции организма. «Как молекулу белка нельзя разделить на части без потери ее специализации, так и ген как единица наследственной информации остается функционально неделимым», при этом генам как единицам хромосомной наследственности функционально соответствуют единицы цитоплазматической наследственности — плазмогены (носители наследственной информации цитоплазатических органелл) (Лобашев и др., 1979, с. 134, 191-192). Поэтому, во-первых, «правильнее было бы расширить понятие генотипа и включить в него как систему хромосомных генов (геном), так и цитоплазматических (плазмон); во-вторых, необходимо различать два явления: 1) наследственность как «свойство генов детерминировать построение специфической белковой молекулы, развитие признака и план строения организма»; 2) наследование как явление, отражающее «закономерности процесса передачи наследственных свойств организма от одного поколения к другому» (Лобашев и др., 1979, с. 4-5, 134).
Впоследствии ген продолжали определять как структурную единицу генетической информации (представленную участком молекулы ДНК, реже РНК), далее неделимую в функциональном отношении (Инге-Вечтомов, 1989). Наконец, Р.Н. Чураев (2006, с. 1277) предложил называть геном «единицу транскрипции — участок молекулы ДНК с примыкающей регуляторной зоной, который кодирует структуру либо одной полипептидной или полирибонуклеотидной цепи, либо несколько цепей, причем в последнем случае участок с необходимостью включает в себя районы, кодирующие начало и конец считывания». Он же предложил и разрабатывает гипотезу об особом классе наследственных единиц — «эпигенов, в которых часть наследственной информации хранится, кодируется и передается потомству вне первичной структуры молекул ДНК генома», т.е. эпиген — «наследственная единица, имеющая не менее двух режимов функционирования подчиненных ей генов и способная сохранять каждый из режимов в поседовательном ряду поколений» (Чураев, 2006, с. 1277). Это приводит к недоразумению: «генетическая система, считающаяся нами управляющей, оказывается управляемой, а управляющей оказывается та, о которой как о системе нам фактически ничего и не известно» (Чадов, 2006, с. 1261).
Между тем, уже из работ генетиков 80-х годов прошлого века было видно, что разработка проблемы гена как центральной проблемы генетики перманентна (Биологический..., 1986) и представления о гене (теория гена) всегда будут отражать «в концентрированной форме уровень развития, достижения и нерешенные проблемы генетики» (Инге-Вечтомов, 1989, с. 370). Указывалось, что «по мере проникновения в молекулярную структуру генетического материала все труднее становится находить в молекулах ДНК границы того, что обозначают понятиями «ген» и «гены» (как наследственные задатки, части генотипа), поскольку «сигналы таких матричных процессов, как транскрипция (на ДНК) и трансляция (на иРНК), не совпадают как по локализации, так и по сочетаниям нуклеотидов»; кроме этого, «наряду со структурными и регуляторными генами, обнаружены участки повторяющихся нуклеотидных последовательностей, функции которых неизвестны, мигрирующие нуклеотидные последовательности (мобильные гены)», а также «псевдогены у эукариот, которые представляют собой копии известных генов, расположенные в других частях генома и лишенные интронов или инактивированные мутациями и поэтому не функционирующие» (Биологический..., 1986, с. 122-123).
В связи с этим, важно отметить, что в 40–50-х годах прошлого века многими отечественными биологами по разным обстоятельствам (не только в силу конъюнктурных соображений) понятие «ген» уже рассматривалось как выдумка формальных генетиков, а передача по наследству приобретаемых (под влиянием окружающей среды) признаков признавалась реальной и объяснимой (см. Иванов, 1960). Затем наступил и продолжается несколько десятилетий этап доминирования прямо противоположных взглядов. Ни в тот, ни в другой периоды развития отечественной биологии по разным причинам не выдвигались концепции, которые могли бы способствовать конструктивному обсуждению и интеграции рациональных элементов альтернативных теорий. Думается, что сейчас в нашей стране такая возможность есть и ее надо использовать для предотвращения в будущем нового этапа официальных абсолютизаций тех или иных положений в генетике и эволюционной биологии. Ведь опасно не инакомыслие, а его административное воплощение.
Поэтому представляется, что только интегрируя рациональные положения, можно корректно разрешить концептуальную коллизию, возникшую вследствие явной односторонности подходов инициаторов «нового синтеза», «эпигенетической теории эволюции» (ЭТЭ), с одной стороны, и сторонников СТЭ — с другой. Современная эволюционная парадигма не должна формироваться путем синтеза положений, только альтернативных СТЭ. Новая парадигма эволюции может быть создана, во-первых, на основе диалектико-материалистического и системно-кибернетического подходов к биологическим объектам и явлениям, во-вторых, путем интеграции рациональных элементов СТЭ и альтернативных взглядов, т.е. концептуальных диалектических пар (например, «борьба за существование–симбиогенез», «эндогенный мутационный процесс–экзогенные экосистемные факторы» и т.д.). Это обусловлено следующими объективными причинами.
Во-первых, с позиций материалистической диалектики любой биологический объект (явление) представляет собой противоречивое единство противоположных сторон (процессов). Такой подход не допускает односторонности взглядов исследователя в вопросах организации, функционирования и эволюции биосистем всех уровней (от молекулярного до биоценотического). Например, при использовании такой методологии неправомерно выделять адаптивные (полезные) и неадаптивные (вредные, нейтральные) признаки, поскольку любой признак биосистемы есть противоречивое единство адаптивных и инадаптивных качеств (Савинов, 1988).
Во-вторых, с позиций кибернетики и биосистемологии (Савинов, 2006) система организменного уровня (СОУ) представляет собой адаптирующуюся самоуправляемую систему. В самом общем виде она может быть представлена единством управляющей (УПС) и исполнительной подсистем (ИПС), образующих вместе контур регулирования (КР) с прямой (от УПС к ИПС) и обратной (от ИПС к УПС) связями. В соответствии с кибернетическим подходом, например, «центральная догма» молекулярной биологии (ДНК→РНК→белок) изначально несостоятельна (Фукс-Киттовский, 1980): она не допускает наличия обратных (хотя и гораздо менее выраженных, чем прямые) связей от белка к РНК и от последней — к ДНК (рис. 1).
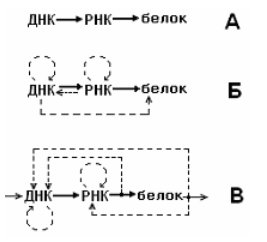
Рис. 1. Схемы, отражающие разные представления об информационных взаимодействиях в системе ДНК–РНК–белок: А — «центральная догма» молекулярной биологии, сформулированная при становлении этой науки; Б — схема, принятая в современной молекулярной биологии (по Инге–Вечтомов, 1989); В — кибернетическая схема, которую можно предложить, рассматривая живую клетку в качестве самоуправляемой биосистемы (по Савинову, 2006)
Представляется, что коллизия между традиционной и эпигенетической концепциями гена разрешается благодаря кибернетической модели элементарной управляющей единицы клетки — элементарной генной единицы (ЭГЕ) (Федоров, 2003). В состав ЭГЕ входит, во-первых, участок ДНК, который соответствует одному гену, кодирует первичную структуру отдельного полипептида и содержит часть регуляторов этого гена; во-вторых, комплекс регуляторов данного гена, находящихся за его пределами; в-третьих, совокупность м-, т-, рРНК, обеспечивающих реализацию генетической информации; в-четвертых, молекулы специфических белков, участвующих в управлении биосинтезом белка. Таким образом, ЭГЕ может быть подразделена на структурную и регуляторную подсистемы: структурная подсистема содержит информацию о структуре синтезируемого полипептида, а регуляторная обеспечивает необходимую степень активности структурной подсистемы (Федоров, 2003). Нетрудно видеть, что такая модель в целом соответствует современным представлениям, во-первых, о ведущей роли ДНК в осуществлении процессов онто- и филогенеза (Корочкин, 2006) и, во-вторых, о координации функций ДНК (посредством ее обратимых модификаций) со стороны продуктов транскрипции и трансляции ДНК и прочих факторов (гистонов, ДНК-метилтрансфераз, морфогенетического поля, механических напряжений в клетках и тканях и др.) (Белоусов, 2006; Ванюшин, 2006; Коряков, 2006; Стегний, 2006). Несомненно, благодаря подобным процессам между управляющими подсистемами «соматических клеток и клеток зародышевого пути существуют межклеточные каналы связи, по которым посредством молекулярных сигналов может передаваться такая информация, что далее она будет наследоваться в последовательном ряду поколений» (Чураев, 2006). Такой постулат современные генетическая и эволюционная парадигмы должны содержать, несмотря на авторитетное противоположное мнение (Корочкин, 2006).
С позиций кибернетики систему организменного уровня (СОУ) в самом общем виде можно представить в форме контура регулирования «генотип–фенотип», в котором УПС соответствует генотип, а ИПС — фенотип (рис. 2).
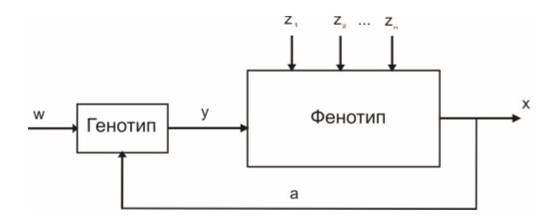
Рис. 2. Схема основного контура регулирования биосистемы организменного уровня (w — задающее воздействие внешней среды, у — регулирующее воздействие, а — обратная связь (информация о выходном сигнале «х»), Z1, Z2... Zn — возмущающие воздействия — «шум») (по Савинову, 2006)
При этом воздействие генотипа на фенотип является прямой связью, а фенотипа на генотип — гораздо менее выраженной обратной связью (Савинов, 2006). Конечно, эта модель выражает лишь суть СОУ («за скобками» остается сложная иерархия многих ее КР). Но даже такая форма системно-диалектического подхода устраняет противопоставление эволюционной роли генотипической изменчивости и экологических факторов, также участвующих в формировании фенотипов, по которым и происходит естественный отбор в системе популяционного уровня (СПУ). Надо лишь учитывать, что, во-первых, в силу эволюционно возрастающей автономизации онтогенеза, филогенетическая роль значимых экологических факторов проявляется гораздо медленнее, чем актуальных генотипических изменений.
Во-вторых, диалектика участия и взаимодействия в онто- и филогенезе генетических и экологических факторов происходит не только вследствие борьбы за существование (комплекса противоречий между СОУ данного биологического вида и внешней средой), но и на основе диалектически альтернативного явления — симбиогенеза (феномена облигатности симбиоза) (Савинов, 2005, 2006). Ведь фактически особь любого вида многоклеточных СОУ (и многих видов одноклеточных СОУ) — это аутоценоз (Савинов, 2005), т.е. особая самоуправляемая система симбионтов (включающая хозяина, его паразитов, мутуалистов, оказывающих управляющее воздействие на хозяина, и вместе выступающих в роли дополнительных УПС СОУ). При таком подходе СПУ — демоценоз (самоуправляемая система аутоценозов), а биологический вид — специоценоз (система демоценозов) (Савинов, 2005, 2006) (рис. 3, 4).
Биологическую эволюцию (биоэволюцию) автор рассматривает как совокупность закономерных, необратимых и уникальных по форме, диалектичных процессов рассеяния и сгущения материи в историческом развитии планеты Земля и сопредельного космоса (Савинов, 2006). Длительное время (порядка 1 млрд. лет) биоэволюция выражалась в необратимом историческом развитии обособившихся от неживой материи разнообразных про- и эукариотических одноклеточных форм. Из них слагались системы популяционно-видового, биоценотического и биосферного уровней. Эти биосистемы положили начало квазициклическим, т.е. «спиралевидным» процессам миграции химических элементов между биотой и земной корой. Однако в ходе эволюции Земли и сопредельного космоса наступил этап, когда для осуществления в возрастающих масштабах процессов рассеяния и сгущения материи «потребовалась» более надежная и высокоэффективная организация биосистем. Некоторые одноклеточные Protozoa, приблизившиеся к состоянию многоклеточности за счет колониальности, несомненно, увеличили свои метаболические возможности по сравнению с прокаритами. Однако и «многоклеточные» простейшие являются биосистемами, организация которых не позволяет вовлекать в их метаболизм эволюционно возрастающие количества веществ, энергии и информации. Проще говоря, эволюционно увеличивать свои объемы и массы могли только сложнее интегрированные и дифференцированные многоклеточные гетеро- и автотрофные биосистемы (Савинов, 2006).
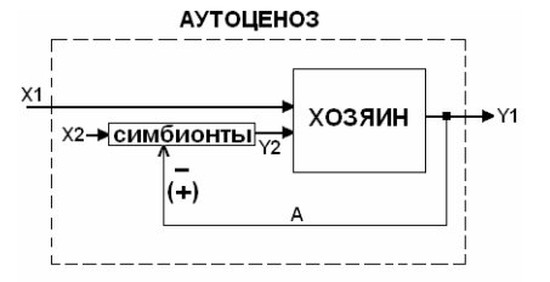
Рис. 3. Общая кибернетическая схема информационных взаимодействий (контур регулирования) в самоуправляемой системе аутоценоза («симбионты–хозяин») (по Савинову, 2005, 2006)
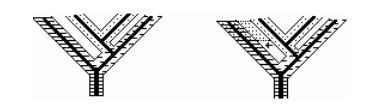
Рис. 4. Схематичное изображение некоторых вариантов филогенеза демо- и специоценозов. Жирными линиями обозначены филумы хозяев, штриховыми и точками — филумы симбионтов (по Савинову, 2006)
Судя по всему, в самом начале своей эволюции прокариоты и одноклеточные эукариоты были рассеяны во всех средах примитивных экосистем биосферы. Среди них не было и не могло быть групп, постоянно закрепленных на каких-либо субстратах или изначально ведущих пассивный образ жизни. Строение клеток этих первичных организмов-биосистем, по-видимому, было обусловлено активным взаимодействием с окружающей средой, выражающемся, очевидно, в различных движениях этих организмов, периодических изменениях формы их клеток и т.п. Однако диалектика эволюции, несомненно, уже в это время породила дивергенцию среди эукариот: одни из них продолжали развивать активный образ жизни, для других предпочтительнее стало формирование пассивных взаимоотношений с окружающей средой. В дальнейшем такая «дилемма» возникала неоднократно и среди представителей групп, которые развивали активный образ жизни. Такие дивергенции всякий раз были обусловлены необходимыми для биосферы режимами обмена веществом, энергией и информацией с окружающим космосом, которые соответствовали характеру сгущения и рассеяния материи в данный период времени под влиянием эндогенных (земных) и экзогенных (космических) процессов во Вселенной. В результате в биосфере сформировались разные типы организации многоклеточных биосистем: такие, среди которых, например, условно можно выделить «растительный», «животный», «грибной», «лишайниковый», а, возможно, и другие типы, в каждом из которых необходимо рассматривать подтипы, адекватные уровню организации соответствующих групп организменного уровня.
Например, сложившийся у высших растений тип организации соответствует их космической роли и весьма наглядно демонстрирует диалектичность космических функций (ассимиляция и диссимиляция) жизни, состоящих, с одной стороны, в сгущении материи, закреплении ее в теле Земли; с другой стороны, в гораздо менее выраженном рассеянии этой материи в сопредельном космическом пространстве. Растения аккумулируют солнечную энергию для синтеза биополимеров из рассеянных простых веществ. Тем самым они способствуют сгущению материи. В результате этих процессов возникли залежи каменного угля и запасы гумуса в современных почвах. Однако одновременно в процессе роста растения удаляют от поверхности Земли химические элементы (материю) и рассеивают часть своей запасенной энергии. «Рекордсменами» такой деятельности, очевидно, являются два вида древесных (высокогорных!) растений — секвойя и секвойядендрон, длина ствола у которых превышает 100 м (!). То, что эти виды — реликты, несомненно, свидетельствует, во-первых, о том, что исторически преобладающей космической функцией живых организмов является сгущение, а не рассеяние материи; во-вторых, это объясняет, почему гигантизм биосистем — относительно кратковременное эволюционное явление (например, в филогенезе динозавров, аммонитов, нуммулитов и др.), обусловленное спецификой отдельных этапов структурно-функциональной эволюции Солнечной системы и Вселенной.
Прогрессирующее обособление аутоценозов от абиотической среды выразилось в автономизации онтогенезов и совершенствовании их саморегуляции (самоуправления). Это обусловило формирование совершенных сенсорных подсистем (неотъемлемых компонентов УПС на организменном уровне), во-первых, позволяющих аутоценозам интегрироваться в демо- и биоценозы за счет физической и химической биокоммуникации, и, во-вторых, минимизировать воздействие нежелательных экологических факторов биотической и абиотической природы. Указанные обстоятельства обеспечивают самосохранение и самовоспроизведение ауто-, демо- и биоценозов, адекватных (благодаря селектогенезу) экологической обстановке в биосфере и сопредельном космическом пространстве. С помощью сенсорных подсистем в аутоценозах корректируется деятельность и структурно-функциональные параметры УПС различных уровней (геномов, нервной системы и т.д.) и ИПС в онтогенезе в соответствии с экосистемными условиями. Это в конечном итоге приводит к филогенетическим изменениям УПС и ИПС различных уровней, т.е. к появлению новых видов аутоценозов. Таким образом, эволюцию биосистем, наряду с другими факторами, обусловливают и долговременные избирательные воздействия исполнительных подсистем (фенотипа) на элементы управляющих подсистем по каналам обратной связи. В свою очередь, эти избирательные воздействия стимулируются влиянием специфических экологических факторов. Однако эффективность таких опосредованных эктогенетических воздействий на молекулярно-генетические и другие управляющие подсистемы невысока. В противном случае нарушатся кибернетические принципы самоуправления и одна из важнейших эволюционных закономерностей — возрастание автономизации процессов индивидуального развития биосистем в ходе филогенеза. При отсутствии коррекции деятельности управляющих подсистем, включая генетические, со стороны управляемых подсистем (в том числе и в зависимости от экологической обстановки) невозможно осуществление онто- и филогенеза. Несомненно, что существенный вклад в данный процесс вносят также мутации, популяционные волны, изоляция и симбиогенез.
Представляется, что высказанные положения могут составлять основу современной парадигмы биологической эволюции.
Литература
Белоусов, Л.В. Морфомеханический аспект эпигенеза / Л.В. Белоусов // Генетика. 2006. Т. 42. № 9. С. 1165–1169.
Биологический энциклопедический словарь. М.: Сов. энциклопедия, 1986. 831 с.
Ванюшин, Б.Ф. Метилирование ДНК и эпигенетика / Ванюшин Б.Ф. // Генетика. 2006. Т. 42. № 9. С. 1186–1199.
Гродницкий, Д.Л. Две теории биологической эволюции / Д.Л. Гродницкий. Саратов: Изд-во «Научная книга», 2002. 160 с.
Иванов, Н.Д. Дарвинизм и теории наследственности / Н.Д. Иванов. М.: Изд-во АН СССР, 1960. 278 с.
Инге-Вечтомов, С.Г. Генетика с основами селекции / С.Г. Инге-Вечтомов. М.: Высш. шк., 1989. 591с.
Иорданский, Н.Н. Эволюция жизни / Н.Н. Иорданский. М.: Academia, 2001. 432 с.
Карпачевский, Л.О. Почвы и социальные науки / Карпачевский Л.О., Зубкова Т.А., Соколов И.А. [ и др.] // Экология и почвы. Т. 5. Пущино: ОНТИ ПНЦ РАН, 2006. С. 5–10.
Корочкин, Л.И. Что такое эпигенетика / Л.И. Корочкин // Генетика. 2006. Т. 42. № 9. С. 1156–1164.
Коряков, Д.Е. Модификация гистонов и регуляция работы хроматина / Д.Е. Коряков // Генетика. 2006. Т. 42. № 9. С. 1170–1185.
Красилов, В.А. Теория эволюции: необходимость нового синтеза / В.А. Красилов // Эволюционные исследования. Макроэволюция. Владивосток: ДВНЦ АН СССР, 1984. С. 4–12.
Красилов, В.А. Нерешенные проблемы теории эволюции / В.А. Красилов. Владивосток: ДВНЦ АН СССР, 1986. 140с.
Лобашев, М.Е., Генетика с основами селекции / М.Е. Лобашев, К.В. Ватти, М.М. Тихомирова. М.: Просвещение, 1979. 304 с.
Назаров, В.И. Учение о макроэволюции: на путях к новому синтезу / В.И. Назаров. М.: Наука, 1991. 288 с.
Назаров, В.И. Эволюция не по Дарвину: смена эволюционной модели / В.И. Назаров. М.: КомКнига, 2005. 520 с.
Савинов, А.Б. К развитию теории адаптациогенеза / А.Б. Савинов // Проблемы макроэволюции. М.: Наука, 1988. С. 153–154.
Савинов, А.Б. Новая популяционная парадигма: популяция как симбиотическая самоуправляемая система / А.Б. Савинов // Вестн. Нижегород. ун-та им. Н.И. Лобачевского. Сер. Биология. 2005б. Вып. 1 (9). С. 181–196. (http://macroevolution.narod.ru/savinov.htm)
Савинов, А.Б. Биосистемология (системные основы теории эволюции и экологии) / А.Б. Савинов. Н. Новгород: Изд-во ННГУ, 2006. 205 с.
Северцов, А.С. Теория эволюции / А.С. Северцов. М.: Владос, 2005. 380 с.
Стегний, В.Н. Эволюционное значение архитектоники хромосом как формы эпигенетического контроля онто- и филогенеза эукариот / В.Н. Стегний // Генетика. 2006. Т. 42. № 9. С. 1215–1224.
Струминский, В.В. Как и зачем возникла жизнь на Земле и других планетах космоса / В.В. Струминский // Вестн. РАН. 1995. Т. 65. № 1. С. 38–51.
Уотсон, Д. Молекулярная биология гена / Д. Уотсон. М.: Мир, 1967. 462 с.
Федоров, В.И. Принципы организации и функционирования живых систем. Ч. 2. Управляющие системы организма / В.И. Федоров. Новосибирск: Изд-во НГТУ, 2003. 142 с.
Фукс-Киттовский, К. Проблемы детерминизма и кибернетики в молекулярной биологии: факты и гипотезы о соотношении между автоматами и живым организмом / К. Фукс-Киттовский. М.: Прогресс, 1980. 375 с.
Хлебосолов, Е.И. Лекции по теории эволюции / Е.И. Хлебосолов. М.: УЦ «Перспектива», 2004. 264 с.
Хохряков, А.П. К вопросу об аксиомах и постулатах современного селекционизма / А.П. Хохряков // Эволюционные исследования. Макроэволюция. Владивосток: ДВНЦ АН СССР, 1984. С. 13–23.
Чадов, Б.Ф. Новый этап в развитии генетики и термин «эпигенетика» / Б.Ф. Чадов // Генетика. 2006. Т. 42. № 9. С. 1261–1275.
Чураев, Р.Н. Эпигенетика: генные и эпигенные сети в онто- и филогенезе / Р.Н. Чураев // Генетика. 2006. Т. 42. № 9. С. 1276–1296.
Яблоков, А.В., Юсуфов, А.Г. Эволюционное учение / А.В. Яблоков, А.Г. Юсуфов. М.: Высш. шк., 2004. 310 с.
ХХI Любищевские чтения. Современные проблемы эволюции. Ульяновск: Ульяновск. гос. пед. ун-т, 2007. С. 60–72
А.Б. Савинов, Проблема новой эволюционной парадигмы (философский, системный и общебиологический аспекты) // «Академия Тринитаризма», М.,
|
|